Síntesis de Palmitato
Lanzadera de citrato: proporciona acetil-CoA y NADPH
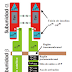
El acetil-CoA es necesario para la síntesis de los ácidos grasos que se lleva a cabo en el citosol, sin embargo, el acetil-CoA se produce en la matriz mitocondrial, por lo tanto existe una ruta para que este salga de la mitocondria y llegue al citosol. Esto ocurre mediante la lanzadera de citrato, el cual se produce a partir de la condensación del acetil-CoA con el oxalacetato, en la primera reacción del ciclo de Krebs, por medio de la citrato sintasa. El citrato entonces, por medio de un transportador de la membrana interna de la mitocondria, es capaz de salir al citosol. En el citosol, por medio de la enzima citrato liasa y la hidrólisis de ATP, se produce acetil-CoA y oxalacetato. De aquí, el acetil-CoA está disponible para la síntesis de ácidos grasos, mientras que el oxalacetato participa en otras reacciones.
El oxalacetato no regresa de inmediato a la mitocondria, primero se reduce por la malato deshidrogenasa citosólica para producir malato, reacción que necesita NADH (proveniente de la reacción catalizada por la gliceraldehído-3-fosfato deshidrogenasa de la ruta glucolítica). El malato después pasa por una descarboxilación oxidativa catalizada por la enzima málica (o malato deshidrogenasa dependiente de NADP) y se produce NADPH que se requiere en la síntesis de ácidos grasos.
El piruvato resultante entra a la mitocondria y se convierte en oxalacetato por medio de la piruvato carboxilasa, el cual puede reaccionar de nuevo con el acetil-CoA para la producción de citrato.
Para la producción de una molécula de palmitato, se requieren 8 moléculas de acetil-CoA, que proporcionarán 8 NADPH de los 14 necesarios para la síntesis del ácido graso. Los 6 NADPH restantes, son proporcionados por la ruta de las pentosas fosfato que se lleva a cabo en el citosol. (Devlin, 2011).
 |
| Figura 1. Lanzadera de citrato. Se utiliza citrato para sacar de la mitocondria el acetil-CoA, necesario en el citosol para la síntesis de los ácidos grasos. Se están gastando 2 ATP para sacar el acetil-CoA y para transferir electrones del NADH al NADPH (útil en la síntesis de los ácidos grasos). (Recuperado de: http://biomodel.uah.es/metab/lanza.htm). |
Formación de malonil-coA
El primer paso en la biosíntesis de los ácidos grasos es la formación de malonil-CoA a partir de acetil-CoA y bicarbonato, catalizada por la acetil-CoA carboxilasa.

Figura 2. Carboxilación del acetil-CoA para la obtención de malonil-CoA catalizada por el complejo acetil-CoA carboxilasa. (Recuperado de: https://quizlet.com/es/333295510/biosintesis-ac-grasos-flash-cards/)
El complejo acetil-CoA carboxilasa consta de 3 enzimas (biotincarboxilasa, proteína portadora de biotina y carboxiltransferasa) y requiere de Mn2+, biotina y ATP para su actividad.
La biotincarboxilasa cataliza la transferencia del grupo carboxil (procedente del bicarbonato) a la biotina. El CO2 activado se une de forma covalente con la biotina. Por su parte, la biotina se encuentra unida a la proteína portadora de biotina mediante un enlace amida con el grupo ε- de la cadena lateral de lisina. El enlace amida con la cadena lateral que une la biotina a la proteína portadora es suficientemente larga y flexible para desplazar la biotina carboxilada a la posición de transferencia de grupo carboxílico a la acetil-CoA en una reacción catalizada por la carboxiltransferasa, produciendo así malonil-CoA.
Además de esta función como punto inicial de la síntesis de ácidos grasos, la malonil-CoA inhibe fuertemente a la carnitina aciltransferasa I sobre la cara externa de la membrana mitocondrial externa. Esto evita un ciclo fútil donde los ácidos grasos experimentan β-oxidación en la mitocondria para fabricar acetil-CoA y dar de nuevo a los ácidos grasos en el citosol. (Devlin, 2011).
Reacciones de la síntesis del palmitato

Figura 3. Reacciones involucradas en la síntesis de palmitato. (Recuperado de: https://www.google.com/url?sa=i&url=https%3A%2F%2Feva.fcien.udelar.edu.uy%2Fpluginfile.php%2F17813%2Fmod_folder%2Fcontent%2F0%2F14.%2520Bios%25C3%25ADntesis%2520%25C3%25A1cidos%2520grasos.pdf%3Fforcedownload%3D1&psig=AOvVaw3ckBHgv14NFNckWUpZh10q&ust=1624417157678000&source=images&cd=vfe&ved=0CAsQjhxqFwoTCNjwkNKfqvECFQAAAAAdAAAAABAJ).
Paso a) "Condensación de Acetil-ACP y Malonil-ACP"
La primera reacción en la síntesis de palmitato es una reacción de condensación mediada por la enzima beta-acil-ACP-sintasa.
El grupo acetilo de la Acetil-ACP se transfiere primero desde la ACP a un tiol de cisteína de la enzima. Después, el grupo carboxilo de la Malonil-ACP activa su carbono metileno para que actúe como nucleófilo y ataque al carbono ceto electrofílico del grupo acetilo. Se desprende entonces una molécula de dióxido de carbono y el ACP del Acetil-ACP, dando lugar al tioéster-beta-cetoacil-ACP. (Devlin, 2011).
Paso b) "Reducción de beta-cetoacil-ACP"
La segunda reacción en la síntesis del palmitato es una reacción de reducción mediada por la enzima beta-cetoacil-ACP-reductasa.
El producto de la primera reacción, el tioéster-beta-cetoacil-ACP, se reduce por medio de la enzima utilizando la reducción de un NADPH + H+; se da la reducción del grupo carbonilo en posición C3 a un grupo hidroxilo. Produciéndose 3-hidroxiacil-ACP. (Devlin, 2011).
Paso c) "Deshidratación de D-3-hidroxiacil-ACP"
La tercera reacción en la síntesis de palmitato es una reacción de deshidratación mediada por la enzima 3-hidroxiacil-ACP-deshidratasa.
Por medio de la enzima se produce la eliminación de una molécula de agua y la producción de un doble enlace tipo trans entre el carbono 3 y el carbono 2. Produciéndose el trans-enoil-ACP. (Devlin, 2011).
Paso d) "Reducción de trans-enoil-ACP"
La cuarta reacción en la síntesis de palmitato es una reacción de reducción mediada por la enzima enoil-ACP-reductasa.
El producto de la reacción tres sufre una reducción para la eliminación del doble enlace (insaturación) utilizando la reducción de un NADPH + H+. Se produce un acil-ACP (butiril-ACP) con dos unidades de carbono más.
El butiril-ACP continuará el segundo ciclo condensándose con un Malonil-ACP, y así por 7 ciclos hasta producir el palmitoil (C16)-S-ACP. (Devlin, 2011).
Paso e) "Hidrolisis de Palmitoil-S-ACP"
La última reacción en la síntesis de palmitato es una reacción de hidrolisis mediada por la enzima tioesterasa.
Por medio del ingreso de una molécula de agua se elimina el ACP y se da formación del palmitato al unirse el grupo hidroxilo en el grupo carbonilo del ácido graso, formando la cabeza polar carboxílica.(Devlin, 2011).
Estequiometría
-La reacción de los siete ciclos para la formación del palmitato es:
Acetil-CoA
+ 7 malonil-CoA + 14NADPH + 14H+↔palmitato + 7CO2 +
14NADP+ + 8CoA-SH + 6H2O
Cabe mencionar que aunque se libera una molécula de agua por cada ciclo, se usa una para la hidrólisis final del palmitoil-S-ACP.
-Tomando en cuenta el gasto de ATP de la síntesis del malonil-coA:
7 Acetil-CoA
+ 7CO2 + 7ATP ↔7 malonil-CoA + 7ADP + 7Pi + 7H+
-Por lo tanto, la reacción global de la conversión de acetil-coA a palmitato es:
8 Acetil-CoA
+ 7ATP + 14NADPH + 14H+↔ palmitato + 14NADP+ + 8CoA-SH +
6H2O + 7ADP+ 7Pi
(Devlin, 2011).
Control de la síntesis de ácidos grasos
El mecanismo de la biosíntesis de los ácidos grasos se logra mediante mensajeros extracelulares como las hormonas. La hormona que participa en la biosíntesis de los ácidos grasos es la insulina, mientras que el glucagón participa en la β-oxidación, es decir, en la degradación de los ácidos grasos.
Los adipocitos son los que se encargan de almacenar y movilizar los triglicéridos para convertirlos en ácidos grasos y transportarlos hacia los órganos, con el objeto de satisfacer sus demandas energéticas. Como mensajeros extracelulares, las hormonas son muy adecuadas para estos papeles reguladores entre los diversos órganos.
La insulina actúa de diversas formas para estimular la síntesis de los ácidos grasos. Uno de sus efectos consiste en estimular la entrada de la glucosa en las células; este efecto aumenta el flujo a través de la glucólisis, es decir, la degradación de la glucosa para producir energía. Por lo tanto, este proceso glucolítico da como producto final al piruvato y mediante la piruvato deshidrogenasa, se convierte en acetil-CoA que se requiere para la síntesis de los ácidos grasos. La activación que induce la insulina sobre la piruvato deshidrogenasa se lleva a cabo a través de un proceso de desfosforilación; la insulina desfosforila a la piruvato deshidrogenasa volviéndola activa y de esta manera convierte el piruvato en acetil-CoA, precursor indispensable para la síntesis de los ácidos grasos. El mecanismo de este efecto de la insulina no está del todo comprendido, pero se deduce que en las fosforilaciones y desfosforilaciones intervienen las fosfatasas; en este caso, la piruvato deshidrogenasa fosfatasa estimulada por los incrementos de las concentraciones de calcio, es la que va a hidrolizar cuando está fosforilada, de esta manera elimina el fosfato de la piruvato deshidrogenasa y la pasa a la forma activa.
Otro mecanismo de regulación es que la mitocondria tiene que estar enviando unidades de acetilo en forma de citrato hacia el citosol, en donde tiene lugar la síntesis de los ácidos grasos.
Una enzima clave que participa en la síntesis de los ácidos grasos es la acetil-CoA carboxilasa. Cuando hemos dejado de comer más de 2 horas, existe una baja concentración de esta enzima; esto refleja probablemente un efecto de control hormonal (glucagón) a través del AMP cíclico. La acetil-CoA carboxilasa es dependiente de la proteína quinasa y tiene que estar desfosforilada para que sea activa; entonces, la fosforilación de la acetil-CoA carboxilasa por la proteína quinasa tiende a inactivar la enzima. En periodo de ayunas, el glucagón inactiva a la enzima acetil-CoA carboxilasa fosforilándola y por lo tanto, no participa en la síntesis de los ácidos grasos. Cabe mencionar que cuando la acetil-CoA carboxilasa está inactiva se encuentra en forma de monómeros; para que participe en la síntesis de los ácidos grasos tiene que estar polimerizada.
El último punto de control tiene que ver con los equivalentes reductores, el NADPH, que proviene del transporte del citrato fuera de la mitocondria y de la ruta de las pentosas fosfato, como se explicó anteriormente.
Cuando se empieza acumular una gran cantidad de NADPH se bloquea la ruta de las pentosas fosfato. Entonces, el segundo mecanismo es la ruta de las pentosas fosfato, se controla mediante la inhibición de la glucosa-6-fosfato deshidrogenasa y de la 6-fosfogluconato deshidrogenasa por el NADPH. Alrededor de un 60% del NADPH para la síntesis de los ácidos grasos procede de esta ruta. (Devlin, 2011).
Referencias bibliográficas
Devlin, T. M. (2011). Textbook of Biochemistry with Clinical Correlations. (7th ed.). USA: John Wiley & Sons, Inc.
Síntesis de ácidos grasos. Recuperado el 20 de junio de 2021, de: https://www.google.com/url?sa=i&url=https%3A%2F%2Feva.fcien.udelar.edu.uy%2Fpluginfile.php%2F17813%2Fmod_folder%2Fcontent%2F0%2F14.%2520Bios%25C3%25ADntesis%2520%25C3%25A1cidos%2520grasos.pdf%3Fforcedownload%3D1&psig=AOvVaw3ckBHgv14NFNckWUpZh10q&ust=1624417157678000&source=images&cd=vfe&ved=0CAsQjhxqFwoTCNjwkNKfqvECFQAAAAAdAAAAABAJ



Es bastante interesante como el palmitato puede jugar un papel muy importante relacionado con la insulina. Desde mi perspectiva, es un blog con información bastante concreta y muy útil por sus mecanismos de reacción si se desea realizar una investigación relacionada con la biosíntesis de este.
ResponderEliminar